|
Л.А.Гаврилов, Н.С. Гаврилова |
"Биология продолжительности жизни" |
|
Назад |
Вперед |
6.8. МОДЕЛЬ НАКОПЛЕНИЯ ДЕФЕКТОВ С ПОСТОЯННОЙ ИНТЕНСИВНОСТЬЮ ПОТОКА ПОВРЕЖДЕНИЙ
Ранее нами были рассмотрены модели, когда интенсивность поступления новых повреждений зависела от степени разрушенности организма (см. разд 6.4-6.7). Так, в модели лавинообразного разрушения (разд. 6.4) интенсивность потока повреждений возрастала по мере увеличения числа дефектов в организме. Напротив, в других моделях (разд. 6.5-6.7) интенсивность потока повреждений уменьшалась по мере уменьшения числа еще неповрежденных элементов (это обусловлено тем, что интенсивность потока повреждений была равна произведению числа еще не поврежденных элементов на постоянную интенсивность их отказа, равную k) Итак, модели с возрастающей и с убывающей интенсивностью потока повреждений нами уже рассмотрены Естественно, для полноты анализа необходимо рассмотреть и третью принципиальную возможность, когда интенсивность потока повреждений, численно равная среднему числу "ударов" (воздействий) в единицу времени, практически не зависит от состояния организма и в среднем постоянна во времени
К подобной модели накопления дефектов могут приводить самые разные представления о разрушении организма. В простейшем случае модель соответствует ситуации, когда на организм воздействует случайный поток внешних ударных нагрузок с постоянной в среднем интенсивностью потока, не зависящей от состояния организма (повреждения экзогенной природы типа космического излучения и т.п.) Возможны, однако, и другие механизмы разрушения, приводящие к данной модели накопления дефектов. В частности, к этой модели можно прийти и путем критического переосмысления предположений, заложенных в описанных ранее моделях (см. разд. 6.5-6 7) Действительно, в этих моделях содержится достаточно спорное утверждение, что смерть организма наступает лишь при отказе всех элементов блока Возможно, что подобная гипотеза и оправдана в ряде случаев для некоторых блоков организма, однако в основном она представляется спорной. Например, трудно представить, чтобы единственная уцелевшая клетка печени (гепатоцит) могла взять на себя функции всей разрушенной печени. Значительно более реалистичной представляется гипотеза, что система исходно содержит огромное число элементов, намного превосходящее то критическое число повреждений, которое приводит к гибели организма. В этом случае мы также приходим к схеме накопления повреждений, причем интенсивность, их потока (равная произведению числа элементов на интенсивность их отказа) оказывается практически постоянной ввиду несоизмеримости числа элементов с допустимым числом повреждений.
Другое достоинство обсуждаемой модели состоит в том. что она позволяет учесть влияние условий жизни на величину критического числа повреждений, несовместимого с жизнью организма. Эта возможность была исключена в ранее рассмотренных моделях (см. разд 6 5-6.7). поскольку гипотеза параллельного соединения элементов предполагала исходную анатомическую заданность их числа для каждого отдельного организма В рамках этих моделей исходное число элементов могло варьировать в зависимости от пола и особенностей популяции, но не могло измениться в ходе жизни организма при изменении условий его жизни В принципе данная особенность моделей хорошо согласуется с тем фактом, что параметры функции Гомперца (/? и а), зависящие o т исходного числа элементов, действительно проявляют историческую стабильность.
Кроме того, варьирование этих параметров в зависимости от пола и особенностей популяции оказывается согласованным так, как это предсказывают математические модели (компенсационный эффект смертности, см. разд. 6.5-6.7). Однако, поскольку историческая стабильность параметров функции Гомперца не является абсолютной. возникает проблема несоответствия моделей параллельного соединения элементов реально наблюдаемым закономерностям.
Ключ к решению данной проблемы состоит в том. чтобы заменить гипотезу параллельного соединения элементов на более близкое к действительности предположение о существовании критического числа повреждений, несовместимого с жизнью организма. В этом случае естественно ожидать, что в более жестких условиях критическое число повреждений, приводящее к гибели, должно быть меньше, чем в более комфортных условиях жизни. В частности, в диких условиях, когда животное лишено ухода и вынуждено само обеспечивать себя пищей, а также спасаться от хищников, первое же серьезное повреждение организма может привести к смерти. Поэтому неудивительно, что смертность многих животных (в частности, птиц) в диких условиях практически не зависит от возраста, что прямо следует из модели одностадийного разрушения организма. Некоторые исследователи на основании подобных фактов отсутствия старения (возрастного увеличения смертности) в диких условиях делают вывод о возможности победить старение путем возврата к естественным "здоровым" условиям жизни без гиподинамии и переедания. Ясно. однако, что борьба с накоплением повреждений путем безжалостного и постоянного уничтожения всех повреждаемых организмов - хотя и эффективный, но малопривлекательный способ борьбы со старением. Напротив, чем большее число повреждений способен накопить организм, оставаясь при этом живым, тем больше будет его продолжительность жизни при прочих равных условиях. Рассуждения подобного рода приводят нас к необходимости углубленного анализа модели накопления дефектов с постоянной интенсивностью потока повреждений и зависящим от условий жизни критическим числом повреждений.
Если интенсивность потока повреждений постоянна и равна k, а организм погибает после накопления n повреждений, то плотность распределения времени жизни организма совпадает с плотностью гамма-функции [Барлоу, Прошан, 1969; 1984]:
![]()
В начальный момент времени это распределение соответствует степенному закону роста интенсивности смертности (закон Вейбулла) с показателем смертности, равным (n - 1).
Принципиально иной результат получается, если учесть исходную поврежденность организмов. Если в начальный момент времени среднее число случайных повреждений в популяции равно
![]() , то вероятность встретить живой организм с
i повреждениями равна:
, то вероятность встретить живой организм с
i повреждениями равна:
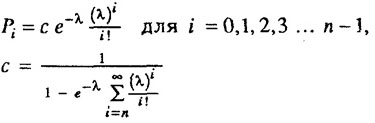
где с - нормирующий множитель, обеспечивающий равенство единице для суммы Pi.
Поскольку смерть организма с i повреждениями наступает после n -1
дополнительных повреждений, то плотность распределения времени
жизни таких организмов равна:
|
|
причем i < n. |
Плотность распределения времени жизни для всей популяции, представляющей смесь организмов с i = 0,1,2, ... n - 1 исходными повреждениями, равна:
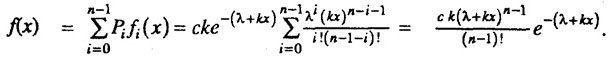
Нетрудно заметить, что в начальный момент времени данная модель также приводит к
биномиальному закону смертности, причем начальный виртуальный возраст популяции
равен ![]() /k. Более подробный анализ модели является излишним, поскольку он формально совпадает с анализом других моделей, проведенным в разделах 6.6-6.7. Отметим, лишь, что в начальный период времени, когда
х <<
/k. Более подробный анализ модели является излишним, поскольку он формально совпадает с анализом других моделей, проведенным в разделах 6.6-6.7. Отметим, лишь, что в начальный период времени, когда
х << ![]() /k, модель приводит к экспоненциальному закону роста интенсивности смертности (закон Гомперца) с показателем степени, равным
k(n-1)/
/k, модель приводит к экспоненциальному закону роста интенсивности смертности (закон Гомперца) с показателем степени, равным
k(n-1)/![]() , и предэкспоненциальным множителем, равным
, и предэкспоненциальным множителем, равным
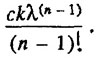 Нетрудно заметить, что обратная связь между этими параметрами Гомперца (компенсационный эффект смертности) может возникнуть как в результате варьирования параметра
Нетрудно заметить, что обратная связь между этими параметрами Гомперца (компенсационный эффект смертности) может возникнуть как в результате варьирования параметра
![]() , (степень исходной поврежденности организмов), так и при варьировании параметра
n (критическое число повреждений, зависящее от жесткости условий существования).
, (степень исходной поврежденности организмов), так и при варьировании параметра
n (критическое число повреждений, зависящее от жесткости условий существования).
Таким образом, основные закономерности смертности могут быть объяснены и в рамках модели накопления дефектов с постоянной интенсивностью потока повреждений, если только организмы исходно содержат значительное число дефектов.
|
Назад |
Вперед |
Дизайн сайта разработан KN Graphics
|
|