|
В.Н. Анисимов |
"Молекулярные и физиологические механизмы старения" |
|
Назад |
Вперед |
1.3. Репродуктивное поведение и эволюция продолжительности жизни
|
За все мы платим звонкою монетой - За чудо жизни и за пламенность утех, За радости любви и что родятся дети. За каждый шаг к вершинам и успех. |
В настоящее время довольно распространена точка зрения, согласно которой генетическая программа развития исчерпывается достижением репродуктивного успеха, то есть рождением потомства и выживанием организма после завершения репродуктивной функции, если и опосредовано геномом, то весьма косвенно (Hayflick, 1998). С эволюционной точки зрения длительный период репродукции дает виду преимущества, тогда как выживание особи после этого периода снижает выживаемость вида (Rose, 1991). Анализ данных о продолжительности жизни и репродуктивной функции у птиц и млекопитающих, живших в неволе (в условиях зоопарка) выявил существенную корреляцию между скоростью возрастного снижения фертильности и скоростью соматического старения (Ricklefs et al., 2003). Ряд недавних публикаций, касающихся связи между возрастом рождения детей и продолжительностью жизни родителей, привлек к этой проблеме пристальное внимание. Было показано, что женщины, которые прожили 100 лет и более, в 4 раза чаще рожали детей после 40 лет, чем те, которые прожили не более 73 лет (Perls et al., 1997). По мнению авторов, поздняя менопауза может быть фактором, способствующим долголетию.
Анализ данных о числе детей и возрасте их родителей в семьях британских аристократов выявил, что эти показатели коррелируют с продолжительностью жизни (Westendorp, Kirkwood, 1998). Оказалось, что среди умерших в молодом возрасте (до 20 лет) дне из каждых трех женщин были бездетными, тогда как среди проживших более 80 лет таких было менее трети. Ранние роды и большое число детей негативно сказывались на продолжительности жизни женщины. Возраст первых родов был наименьшим у умерших рано и наибольшим у проживших более 80 лет. Шансы дожить до 100 лет имели больше те женщины, которые родили первенца после 40 лет. Интересно, что и мужья жили дольше, если число произведенных ими детей было не слишком велико. Продолжительность жизни дочерей больше коррелирует с продолжительностью жизни матери, чем отца, тогда как у сыновей эта зависимость значительно менее выражена и не коррелирует с полом родителей. Авторы делают вывод о том, что, несмотря на некоторую наследуемость продолжительности жизни (Gavrilova, Gavrilov, 1998; Finch, Ruvkun, 2001), каждый человек должен выбирать между долголетием и продолжением рода (рис. 2 и 3). У дрозофил, которым не позволяли размножаться, продолжительность жизни увеличивалась.
Проанализировав данные о гетерозиготности генома у 77 зоологических и 30 ботанических видов, Ю. П. Алтухов (1996) установил, что она с большой степенью достоверности коррелирует со скоростью полового созревания и отрицательно - с продолжительностью жизни. Чем больше индивидуальная гетерозиготность, тем большие энергетические закаты организма приходятся на этот период онтогенеза, выше темпы развития, раньше наступает половозрелость и возраст первой репродукции, и соответственно короче оказывается жизнь. Подчеркивается, что долгожительство человека, как и у других биологических видов, определяется высокой индивидуальной гомозиготностью.
Другим аспектом проблемы продолжительности жизни потомства является оценка генетического груза мутаций, полученных родителями в процессе старения. Частота мутаций в половых клетках мужчин много выше, чем у женщин, и пропорциональна возрасту. В случае отцовства в позднем возрасте потомство подвергается большему риску генетических нарушений и соответственно риску иметь меньшую продолжительность жизни (Crow, 1997). Особенно это касается дочерей старых отцов. Продолжительность жизни у таких дочерей явно короче. В сперматогенных клетках старых мышей частота мутаций была увеличена по сравнению с таковой у молодых самцов или особей среднего возраста (Walter et al., 1998). Однако анализ данных по 320 столетним, рожденным во Франции между 1875 и 1890 гг. и умершим между 1990-1999 гг., показал, что возраст отцов на момент рождения лиц, достигших ста лет, не отличался от возраста отцов контрольной популяции, причем и у дочерей (Robine et al., 2003).
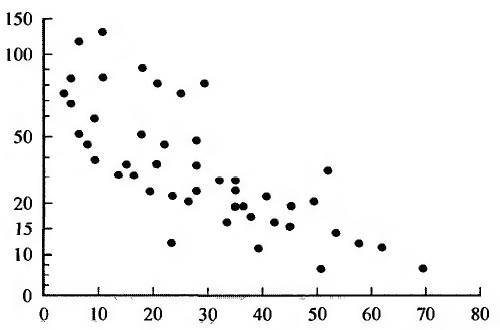
Рис. 2. Зависимость между репродуктивным потенциалом и долгожительством у 47 видов млекопитающих.
По оси абсцисс - продолжительность жизни, годы; по оси ординат - потенциальное число потомков, log.
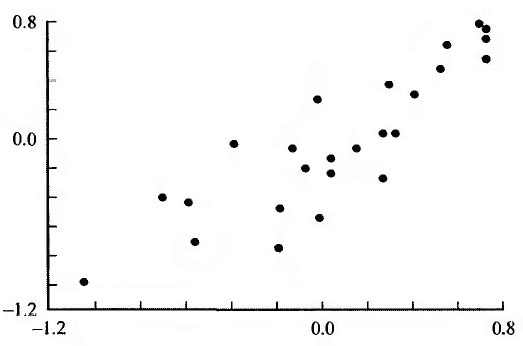
Рис. 3. Зависимость между относительным возрастом первых родов и продолжительностью жизни.
По оси абсцисс - относительная продолжительность жизни; по оси ординат - относительный возраст первых родов.
В опытах на крысах не наблюдалось различий в продолжительности жизни потомства, произведенного молодыми (3 мес.) самками и самцами и молодыми самками и старыми (27 мес.) самцами, однако потомство старых самцов было значительно более чувствительно к канцерогенному действию N-нитрозометилмочевины (Anisimov, Gvardina, 1995), что свидетельствует в пользу точки зрения о накоплении с возрастом генетических повреждений в мужских половых клетках. В эпидемиологических исследованиях установлен повышенный риск развития опухолей нервной системы, ретинобластомы и острого лимфобластного лейкоза у потомства старых родителей, особенно отцов (Hemminki, Kyyronen, 1999; Dockerty et al., 2001).
|
Назад |
Вперед |
Дизайн сайта разработан KN Graphics
|
|